 Nonalcoholic steatohepatitis (NASH) is associated with a chronic proinflammatory state of the liver1-3,10. The clinical pathogenesis of NASH progresses from a steatotic/intralobular inflammatory/hepatocellular ballooning state to progressing levels of fibrosis and in later stages, an increased incidence of cirrhosis and/or hepatocellular carcinoma8-13. The etiology underlying this progression remains unknown, nor are there currently any approved therapies other than lifestyle modifications aimed at modulating or mitigating NASH11,12.
Nonalcoholic steatohepatitis (NASH) is associated with a chronic proinflammatory state of the liver1-3,10. The clinical pathogenesis of NASH progresses from a steatotic/intralobular inflammatory/hepatocellular ballooning state to progressing levels of fibrosis and in later stages, an increased incidence of cirrhosis and/or hepatocellular carcinoma8-13. The etiology underlying this progression remains unknown, nor are there currently any approved therapies other than lifestyle modifications aimed at modulating or mitigating NASH11,12.
Identifying New NASH Drug Targets
Han and colleagues sought to identify and define a key pathway responsible for the chronic activation of hepatic proinflammatory immune responses that define the NASH-associated hepatic environment. By understanding the regulators of this immune homeostatic pathway, the authors identify targets that may offer therapeutic potential against NASH.Macrophages and NASH
The critical role of macrophages in the hepatic proinflammatory state of NASH, specifically Kupffer cells (KC), has supported further investigation into the pathways that regulate their function1,2,4,5,7,8. Macrophages have two functional states: an M1 proinflammatory and a M2 anti-inflammatory polarity1,2,8. The M1 polarized macrophages exacerbate the NASH inflammatory state and result in liver injury via the production of proinflammatory cytokines such as tumor necrosis factor α (TNFα). In contrast, M2 macrophages maintain an anti-inflammatory state by suppression of M1 macrophage activation, apoptosis of M1 macrophages, and the production of anti-inflammatory cytokines such as IL-101,2,6.The M1/M2 Polarity Switch
The M1/M2 polarization of macrophages is associated with the progression of NASH with patients showing higher M2 levels with steatosis vs. patients with severe steatohepatitis1. Recent studies have shown that by inducing a polarity switch in hepatic macrophages from M1 to M2, it can resolve a high fat diet-induced animal model of NASH1,2,4,5.Based on their previous work, the authors focused on understanding the pathway governing the polarity switch of M1 to M2 in hepatic macrophages, specifically the involvement of the nuclear receptor retinoic-acid-related orphan receptor α (RORα; NR1F1) in KCs. RORα is a ligand-dependent transcriptional factor that controls an array of target genes associated with both lipid metabolism and inflammation1,2.
ROR Activation in Animal Models
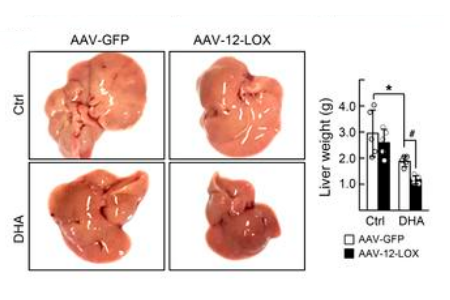
Depicted are a representative sample of the livers and corresponding
liver weights of male C57BL/6 mice exposed to HFD for 16 weeks. Animals
were given either AAV-GFP control or AAV-LOX-12 (5x106 vg; i.v.)
at approximately 13 weeks of diet exposure and were then subsequently
treated for the remaining 3 weeks with DHA (1mg/kg; i.p.; q.d.) or vehicle.
Source: Han et al. JCI 20191
NASH pathogenesis was exacerbated in a RORα Knockout (KO) mouse model. The RORα KO showed significant alterations in polarity switching with M1 macrophage levels elevated compared to M2. This RORα- dependent exacerbation of NASH was due to effects on the macrophage polarity ratio and not due to alterations in overall macrophage populations1.
Further Investigation of RORα in NASH
Due to the newfound importance of RORα in the development of NASH, the authors sought to identify the components of hepatic milieu that effect the function of this receptor. They narrowed their focus to polyunsaturated fatty acids (PUFAs) and their metabolites due to the altered ratio of PUFAs to saturated fatty acids (high saturated and low PUFAs) in NASH patients1,13. Additionally, FAs are also known to modulate macrophage polarity switch and are thought to be an essential source of M2 activation with docosahexaenoic acid (DHA) being an ideal candidate1.The authors determined that a metabolite of DHA, maresin-1 (MaR1; unlike DHA) was an endogenous agonistic ligand for RORα in hepatic macrophages.
In Vitro Studies of RORα
In vitro studies showed that MaR1 activation of RORα resulted in a M2 polarity switch (determined via M1/M2 cell marker ratios and M2 signature gene expression levels) in liver macrophages that was both dose and time-dependent1. MaR1 also increased the expression (both mRNA and protein levels) of RORα in liver macrophages1. The effects of MaR1 to induce a M1 to M2 polarity switch in hepatic macrophages required the presence of RORα1.Further In Vivo Studies
“...these results suggest that the MaR1/RORa/12-LOX autoregulatory circuit could offer potential therapeutic strategies for curing NASH.”
Additionally, in vivo studies showed that Mar1 could inhibit the progression of NASH in an HFD mouse model only in the presence of RORα1. Mar1 increased the number of M2 hepatic macrophages in HFD fed flox control mice, whereas, the levels were unaltered in the HFD fed RORαKO mice1. Similar to RORα, Mar1 expression levels were significantly depressed in both an HFD and methionine choline-deficient (MCD) diet exposed mouse models when compared to controls1. Restoration of Mar1 levels could be addressed via DHA administration but only in the presence of RORα.
–Han et al.
Furthermore, clarity was gained on the feedback between Mar1 and RORα when a synthetic agonist to RORα increased Mar1 levels, while a KO of RORα decreased them. The function of RORα is associated with the levels of Mar1 via the effects of RORα on the enzyme 12-lipoxygenase (12-LOX), which is involved in the biosynthesis of Mar11.
Han and colleagues showed that RORα increases levels of Mar1 through induction in the expression of 12-LOX. In vitro work showed hepatic macrophages had significantly elevated levels of 12-LOX when compared to other types of macrophages or hepatocytes. Furthermore, in vivo studies demonstrated that RORαKO mice had reduced levels of 12-LOX compared to control. Activation of RORα via synthetic agonist, endogenous agonist, or infection of AAV-RORα increased 12-LOX levels whereas knockdown of RORα decreased them suggesting 12-LOX is a downstream target of RORα.
Regulating the Protective Effects of DHA
 to anti-inflammatory (M2) phenotype.")
A schematic of the MaR1-RORα-12-LOX autoregulatory circuit. Targeting of this
circuit could provide a therapeutic response in NASH via its resulting polarity
switch in liver macrophages from an inflammatory (M1) to anti-inflammatory (M2)
phenotype. Source: Han et al. JCI 20191
In contrast, the symptoms of HFD NASH mice significantly improved when given AAV-12-LOX and DHA (reduction of proinflammatory cytokines, lipids, and histopathological progression) with Mar1 and M2 macrophages levels elevated compared to controls1. The potential translation of 12-LOX as a downstream target extended to the clinic, with mRNA levels of 12-LOX significantly lower in NASH patients compared to healthy controls1. Furthermore, RORα and 12-LOX expression levels in humans show a positive correlation, both are decreased in hepatic macrophages of patients with liver fibrosis compared to controls and are induced by Mar1 or a synthetic agonist to RORα in vitro1.

